Batteri
Funzioni e struttura dei batteri
I Batteri sono microrganismi unicellulari di tipo procariotico. Nella classificazione degli organismi viventi i Batteri o Eubatteri rappresentano uno dei tre domini in cui sono riconducibili tutte le forme di vita attualmente viventi e quelle ormai estinte.
Gli altri due domini sono quello degli Archea, organismi di tipo procariotico come i Batteri, e quello degli Eucarioti.
Gli Archea o Archeobatteri sono microorganismi diffusi soprattutto in ambienti caratterizzati da condizioni estreme di temperatura, pH, salinità, ma se ne trovano molti anche nel suolo, negli ambienti acquatici e in altri organismi viventi, come nello stomaco dei bovini.

Albero filogenetico che descrive le relazioni evolutive tra gli esseri viventi. Tutti gli organismi viventi appartengono a uno dei tre domini Bacteria, Archaea e Eucarya. I primi due costituiscono il gruppo dei procarioti e condividono con gli eucarioti l'origine da un progenitore comune a partire dal quale si sono differenziate le tre principali linee evolutive che hanno dato origine alle forme di vita che noi conosciamo.
Distribuzione dei batteri e utilità per l'uomo
Essendo visibili solo con l'ausilio di un microscopio, è difficile apprezzare come tutti i procarioti siano le forme di vita più abbondanti sulla Terra, sia in termini di biomassa che di numero di specie.
Attualmente sono note circa 3000 specie di organismi procariotici, un numero che si pensa sia meno dell'1% delle specie realmente esistenti.
Sebbene alcuni batteri siano responsabili di malattie anche gravi negli animali e nelle piante, la stragrande maggioranza è del tutto innocua o addirittura benefica.
I batteri si trovano in qualsiasi ambiente, terrestre, acquatico e aereo.
Molti vivono in simbiosi con altri organismi viventi, come i batteri che costituiscono la flora intestinale nell'uomo, coinvolti in vari processi: digestione, produzione di vitamine, difesa immunitaria nei confronti di microrganismi patogeni.
Anche la capacità dei ruminanti di digerire la cellulosa è dovuta ad enzimi prodotti da batteri che si trovano in un particolare organo digestivo, il rumine.
L'agricoltura si basa in gran parte sull'attività microbica.
Le leguminose, ad esempio, vivono in stretta associazione con una specie batterica in grado di fissare l'azoto atmosferico (N2) in NH3, utilizzabile per la sintesi di proteine e altri composti azotati entrando così nella catena alimentare.
Oltre che dell'azoto, i batteri giocano un ruolo fondamentale anche nei cicli biogeochimici di carbonio e zolfo.
Da sempre l'uomo ha sfruttato i batteri in campo alimentare.
I batteri lattici come quelli del genere Lactobacillus, Lactococcus e Streptococcus sono utili nella produzione di yogurt, formaggi, burro e latticini vari.
I batteri lattici e acido-acetici sono utilizzati nella preparazione di prodotti sotto aceto.
Il sapore e l'aroma di prodotti come salami e prosciutti affumicati dipende dai processi fermentativi di alcuni batteri.
Alcuni batteri sono in grado di demolire sostanze inquinanti e trovano impiego nei processi di biorisanamento.
In campo sanitario si sfruttano alcune specie batteriche perché produttrici di antibiotici, usati nel trattamento di infezioni causate da batteri patogeni.
In ultimo, ma non meno importante, va ricordato l'ampio utilizzo dei batteri nelle biotecnologie.
Le caratteristiche della cellula batterica
I batteri hanno cellule di tipo procariotico, quindi mancano di involucro nucleare a protezione del materiale genetico, che si trova nel citoplasma, e non hanno organuli e citoscheletro, caratteristiche tipiche delle cellule eucariotiche.
I batteri sono più piccoli delle cellule eucariotiche.
Le dimensioni cellulari variano da un minimo di 0,2 µm a un massimo di 50-80 µm di diametro.
La morfologia della cellula è variabile. I batteri di forma sferica o ovoidale sono chiamati cocchi, quelli di forma cilindrica bacilli, mentre gli spirilli hanno forma spiraliforme.
Sono frequenti anche morfologie più complesse, come quella delle spirochete, dei batteri peduncolati o dei vibrioni.
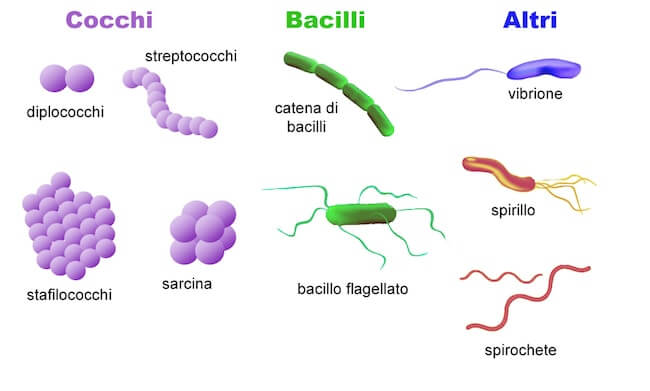
Le forme più comuni riscontrabili nei batteri.
Costituenti citoplasmatici
Nel citoplasma dei batteri sono sempre presenti ribosomi per la sintesi proteica e il DNA.
Il DNA occupa una regione del citoplasma detta nucleoide, che si colora leggermente.
In Escherichia coli, il batterio più studiato, è presente una sola molecola di DNA circolare, detta cromosoma.
Questa organizzazione del DNA è stata usata come modello per descrivere il genoma di tutte le cellule procariotiche, ma la conoscenza sempre più approfondita in questo campo ha portato alla scoperta anche di genomi lineari e di casi in cui sono presenti più copie di DNA circolare o lineare.
Il DNA batterico è ancorato in alcuni punti a delle introflessioni della membrana plasmatica, dette mesosomi, che svolgono un ruolo fondamentale nella duplicazione del DNA e nella ripartizione delle molecole di DNA alle cellule figlie durante la scissione binaria.
I ribosomi hanno dimensioni 70S, più piccoli quindi di quelli eucariotici, di dimensioni 80S.
La subunità minore è pari a 30S e quella maggiore a 50S.
Le piccole differenze riscontrabili tra i ribosomi di procarioti ed eucarioti sono alla base dell'azione selettiva di alcuni antibiotici che, riconoscendo in modo specifico le componenti proteiche o di RNA ribosomiale, bloccano la sintesi proteica e causano la morte delle cellule batteriche, senza interferire con i meccanismi traduzionali delle cellule eucariotiche.
I plasmidi
Nei batteri e nei procarioti in genere, oltre al cromosoma possono essere presenti molecole di DNA che conferiscono caratteristiche accessorie e si replicano indipendentemente dal cromosoma batterico, i plasmidi.
Si tratta generalmente di molecole circolari, che non recano funzioni essenziali alla sopravvivenza e al mantenimento della cellula batterica, ma contengono geni che possono conferire un vantaggio per la crescita in particolari condizioni.
Tra i più noti si ricordano i plasmidi R, che recano geni responsabili della resistenza agli antibiotici, funzione che diventa fondamentale per la sopravvivenza in caso di esposizione a questi agenti antimicrobici.
Alcuni plasmidi consentono di demolire sostanze inquinanti o produrre sostanze responsabili della patogenicità dei batteri.
Nel genere Rhizobium, la capacità di formare noduli nelle radici delle leguminose con cui vivono in simbiosi e fissare l'azoto è dipendente da funzioni codificate da plasmidi.
Alcuni plasmidi, come il plasmide F di Escherichia coli, contengono geni che ne determinano la possibilità di essere trasferiti in altre cellule, mediante il processo di coniugazione, un meccanismo di scambio genico orizzontale che consente di aumentare la variabilità genetica e acquisire capacità nuove utili alla sopravvivenza.

Trasferimento di un plasmide mediante coniugazione. I plasmidi che sono in grado di realizzare questo processo prima si duplicano creando due copie, una delle quali rimane nella cellula donatrice, l'altra viene trasferita alla cellula ricevente. Il contatto cellula-cellula e il trasferimento del plasmide possono avvenire tramite pilo sessuale, come si può osservare nell'immagine al microscopio elettronico sulla destra.
Altre inclusioni e strutture citoplasmatiche
All'interno dei batteri possono essere visibili al microscopio ottico granuli e altre inclusioni, spesso delimitati da un monostrato di membrana di natura lipidica.
La maggior parte delle inclusioni contiene sostanze di riserva come acido poli-β-idrossibutirrico o glicogeno.
Sono presenti anche granuli di polifosfati, per la sintesi di acidi nucleici e fosfolipidi, granuli di zolfo elementare e magnetosomi, contenenti magnetite (Fe3O4), un composto che guida i movimenti cellulari lungo le linee di campi geomagnetici.
In molti batteri acquatici, come i cianobatteri o alghe azzurre, sono presenti vescicole ripiene di gas in grado di garantirne il galleggiamento.
Alcuni batteri sono in grado di formare al loro interno delle endospore, strutture altamente resistenti alla disidratazione, alle radiazioni, a vari detergenti chimici e al calore, che garantiscono la sopravvivenza del batterio in condizioni avverse e ne consentono la diffusione tramite vento, acqua o l'apparato gastrointestinale degli animali.
La membrana citoplasmatica
Come tutte le cellule, anche i batteri sono delimitati da una membrana costituita da un doppio strato di fosfolipidi che costituiscono un mosaico fluido dove le proteine ad essi associati sono dotati di una discreta mobilità.
A differenza delle cellule eucariotiche, la membrana plasmatica non contiene steroli (come il colesterolo nelle cellule animali), ma molecole simili chiamate opanoidi.
Inoltre, gli acidi grassi polinsaturi sono piuttosto rari.
Le proteine svolgono varie funzioni, tra cui riconoscimento di molecole provenienti dall'esterno, attività enzimatica e di trasporto.
La parete cellulare
Tutti i procarioti sono delimitati da una parete cellulare che conferisce forma e rigidità alla cellula e la protegge da alcuni agenti fisici e chimici.
In assenza di parete, i batteri morirebbero in seguito a disidratazione o ingresso di acqua fino a causarne la lisi.
Non presentano parete i micoplasmi, un gruppo di microrganismi patogeni di piante e animali, dotati invece di una membrana plasmatica più resistente.
La parete dei Batteri è costituita di un polimero detto peptidoglicano, un eteropolisaccaride ottenuto dalla ripetizione di N-acetilglucosamina e acido N-acetilmuramico uniti da un legame β 1-4 glicosidico.
Il legame β 1-4 glicosidico è riconosciuto e scisso dal lisozima, un enzima presente nella saliva, nelle lacrime, nel bianco d'uovo e in diversi fluidi animali.
All'acido N-acetilmuramico è legato un gruppo di aminoacidi, tra cui ne figurano alcuni in configurazione D.
Le catene di peptidoglicano sono tenute insieme da legami peptidici, detti legami crociati, tra i gruppi di aminoacidi.
La formazione di questi legami è catalizzata dagli enzimi transpeptidasi, con la cui attività interferiscono gli antibiotici β-lattamici, come le penicilline.
La mancata formazione dei legami crociati indebolisce la parete e stimola il batterio a produrre degli enzimi che la degradano completamente.
Il batterio rimasto senza parete muore per lisi osmotica.
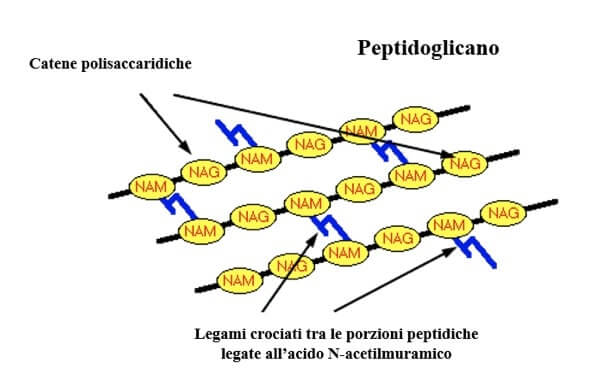
Il peptidoglicano è l'eteropolisaccaride che costituisce la parete batterica. La parte polisaccaridica è data dalla ripetizione di N-acetilglucosamina (NAG) e acido N-acetilmuramico (NAM). A quest'ultimo è legato un gruppo di aminoacidi che forma legami crociati con gli aminoacidi della catena adiacente.
Gram positivi e Gram negativi
Sulla base della struttura della parete i Batteri si dividono in due grandi gruppi: Gram positivi e Gram negativi.
La distinzione dipende dalla diversa colorazione assunta utilizzando la tecnica messa a punto dal microbiologo danese Hans Christian Gram nel 1884.
I Gram positivi si colorano di viola, i Gram negativi di rosa.
La tecnica prevede l'utilizzo di un complesso insolubile di cristal-violetto, che rimane intrappolato nei Gram positivi, rimosso invece dall'alcol, usato nella seconda fase della colorazione, nei Gram negativi.
I Gram positivi hanno una parete più spessa, costituita di più strati di peptidoglicano che la rendono impermeabile all'alcol, lasciando il complesso di cristal-violetto nella cellula.
La parete dei Gram negativi è più sottile ed è inoltre presente un'ulteriore membrana, detta membrana esterna, permeabile al solvente che, data la parete più sottile, riesce a penetrare nella cellula e a rimuovere il complesso di cristal-violetto.
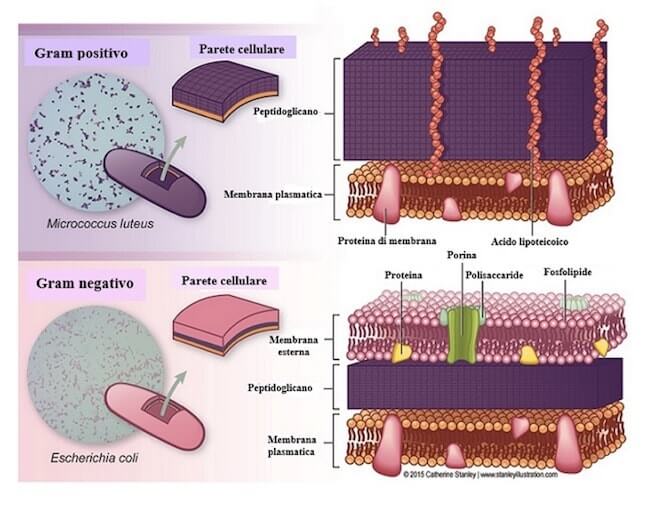
Le differenze della parete cellulare dei batteri Gram positivi e Gram negativi, da cui dipende la diversa colorazione, viola per i primi, rosa per i secondi. Gli acidi lipoteicoici sono polisaccaridi tipici della parete dei Gram positivi a cui conferiscono, data l'elevata densità di cariche negative, la capacità di legame ioni positivi, come Ca2+ e Mg2+.
La membrana esterna dei Batteri Gram negativi
La membrana esterna, in aggiunta al sottile strato di peptidoglicano presente nei batteri Gram negativi, contiene un doppio strato fosfolipidico, come la membrana plasmatica, ma anche numerosi polisaccaridi, legati a diversi lipidi con cui formano dei complessi lipopolisaccaridici.
Per tale ragione la membrana esterna è anche chiamata lipopolisaccaride o LPS.
La membrana esterna presenta una maggiore permeabilità a piccole molecole rispetto a quella interna.
Ciò è dovuto alla presenza di proteine-canale chiamate porine, che consentono il passaggio di varie sostanze idrofiliche.
La regione che separa la membrana esterna dalla parete è detta periplasma, dove si trovano enzimi, proteine di trasporto e proteine con funzione recettoriale.
La membrana esterna non ha solo un ruolo strutturale in quanto è coinvolta nei meccanismi di patogenicità di alcuni Gram negativi, come i membri dei generi Salmonella, Shigella ed Escherichia, responsabili di disordini gastrointestinali (vomito, diarrea) derivati ad esempio da infezioni alimentari.
La tossicità si deve a una porzione lipidica dell'LPS, che è un'endotossina, cioè un componente strutturale della cellula con proprietà tossiche.
Strutture di superficie della cellula batterica
Sulla superficie di alcuni batteri è possibile trovare delle strutture filiformi di natura proteica: fimbrie, pili e flagelli.
Le fimbrie consentono l'adesione alle superfici come i tessuti animali nel caso di batteri patogeni.
I pili, simili alle fimbrie, ma meno numerosi e più lunghi, possono essere coinvolti nell'adesione alle superfici, ma sono principalmente utilizzati nella coniugazione (pili sessuali). I pili, inoltre, sono utilizzati come recettori da parte di alcuni virus.
I flagelli sono appendici con struttura elicoidale ottenuti dall'assemblaggio di numerose subunità della proteina flagellina.
Il movimento rotatorio dei flagelli assicura la motilità della maggior parte dei batteri che vivono in ambienti acquatici.
La disposizione dei flagelli è varia. Se sono attaccati a uno o a entrambi i poli, la distribuzione è detta polare; lofotrica se è presente un ciuffo si flagelli a un'estremità; peritrica se sono distribuiti su tutta la cellula.
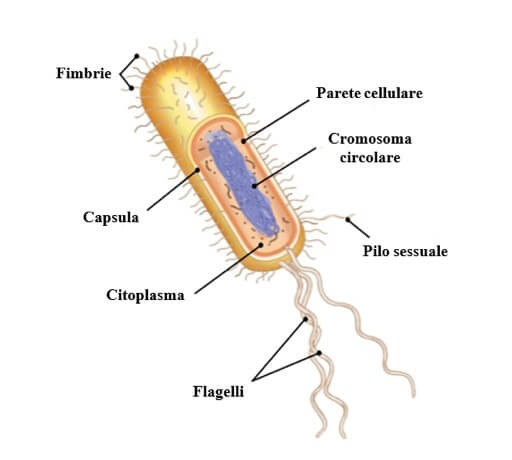
Sulla superficie batterica possono trovarsi appendici proteiche con varie funzioni: fimbrie, pili e flagelli.
Ulteriori rivestimenti esterni
Alcuni batteri secernono sulla superficie esterna sostanze polisaccaridiche che formano degli ulteriori rivestimenti con varie funzioni.
Se hanno una struttura rigida e impermeabile a varie sostanze prendono il nome di capsule, mentre nel caso di un'organizzazione meno compatta si parla di strati mucosi.
Questi involucri sono implicati nell'adesione alle superfici, compresi i tessuti animali, e nei meccanismi di patogenicità in quanto in alcuni casi impediscono il riconoscimento da parte delle cellule fagocitiche dell'organismo ospite.
Possono inoltre contribuire alla protezione dall'essiccamento data la capacità di richiamare grandi quantità di acqua.
Insieme alle fimbrie, la capsula gioca un ruolo fondamentale nella formazione di biofilm, pellicole che si formano sulle superfici più disparate (ad esempio rocce e tessuti animali), dove un gran numero di batteri sono immersi e circondati da una matrice polisaccaridica, che regola i processi di scambio di sostanze nutritive e di eliminazione di quelle di rifiuto e, soprattutto, conferisce resistenza a svariati agenti chimici, come gli antibiotici e protegge dal dilavamento nel caso in cui siano esposti a flussi di fluidi.
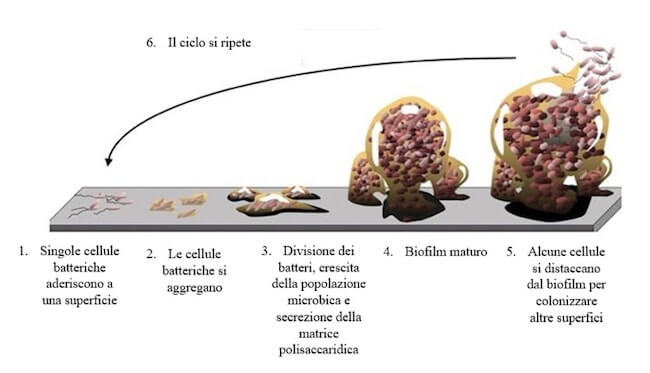
Il biofilm rappresenta la forma più comune in cui più cellule batteriche si aggregano e formano pellicole adese alle superfici.
Riproduzione e meccanismi di variabilità genica
I batteri si riproducono asessualmente per scissione binaria, mediante la quale da una cellula madre si ottengono cellule figlie geneticamente identiche, a meno di eventuali mutazioni insorte durante la duplicazione del DNA.
Grazie alla scissione binaria il numero di cellule di una popolazione aumenta considerevolmente in poco tempo, processo definito crescita.
La crescita di una popolazione batterica si realizza in tempi diversi a seconda della specie e, soprattutto, della disponibilità di nutrienti.
In condizioni ottimali, il processo di divisione in Escherichia coli si completa in circa 20 minuti, ma sono noti batteri che si riproducono anche più velocemente.
La riproduzione asessuata non consente di realizzare ricombinazioni geniche che potrebbero aumentare la variabilità della popolazione batterica, da cui dipendono maggiori probabilità di sopravvivenza nel caso in cui le condizioni ambientali varino.
Tuttavia, nei procarioti sono noti meccanismi di scambio genico tra cellule non necessariamente appartenenti alla stessa specie ed evolutivamente correlate.
Questi processi, detti di trasferimento genico orizzontale, sono nel complesso definiti anche come meccanismi di parasessualità in quanto concorrono a conferire proprietà nuove ai batteri che ne sono coinvolti, come accade nel caso degli organismi che si riproducono sessualmente.
Oltre alla già citata coniugazione, il trasferimento genico orizzontale può avvenire anche mediante trasformazione e trasduzione.
La trasformazione consiste nell'assunzione di frammenti di DNA rilasciati da cellule morte nelle vicinanze.
La trasduzione prevede il trasferimento di regioni di DNA da un batterio a un altro tramite un fago.
Durante la fase di impacchettamento dei nuovi virioni in una cellula ospite, può accadere che nella testa venga inserito oltre al DNA fagico anche qualche tratto di DNA batterico.
Quando questi fagi verranno rilasciati e infetteranno altre cellule potranno introdurre in esse anche il DNA batterico, che eventualmente conferirà loro caratteristiche nuove in grado di ottimizzarne la sopravvivenza.
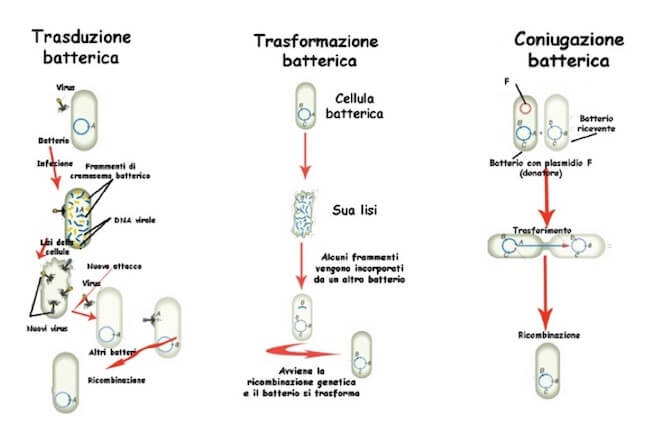
Panoramica dei meccanismi di trasferimento genico orizzontale nei procarioti.
Metabolismo e adattamento alle condizioni ambientali
I batteri possono essere autotrofi o eterotrofi.
Tra gli autotrofi, la fonte energetica utilizzata può essere la luce (fotoautotrofi) o l'energia liberata dall'ossidazione di composti inorganici, come H2, H2S e NH3 (litoautotrofi).
Riguardo l'ossigeno, i batteri aerobi utilizzano questo gas nel loro metabolismo e alcuni sono in grado di tollerarlo anche a concentrazioni maggiori di quella atmosferica.
I microaerofili possono utilizzare O2 solo a livelli inferiori di quelli atmosferici e gli aerobi facoltativi possono crescere sia in sua presenza che in sua assenza.
I batteri che non possono utilizzare ossigeno nel loro metabolismo sono detti anaerobi, i quali possono essere aerotolleranti se possono comunque crescere in sua presenza, obbligati se ne sono invece inibiti o uccisi.
I batteri si differenziano notevolmente per il range di temperatura di cui necessitano per la sopravvivenza.
A questo proposito si distinguono:
- batteri mesofili, con temperature ottimali a valori medi;
- batteri termofili capaci di vivere a temperature superiori ai 45 °C;
- batteri ipertermofili, il cui optimum di temperatura è superiore agli 80 °C.
La maggior parte dei batteri vive in ambienti caratterizzati da pH compresi tra 5 e 9, ma sono anche noti batteri acidofili, capaci di vivere anche in presenza di pH inferiore a 2, e basofili, adattati a condizioni di pH superiori a 9.
Molta variabilità esiste anche per la concentrazione di NaCl che deve essere presente nell'ambiente esterno per svolgere in modo ottimale le funzioni fisiologiche.
Sono definiti alofili i batteri che necessitano di concentrazioni di NaCl comprese tra 1% e 15%, alotolleranti quelli che non hanno particolari esigenze in questo senso.
Patogenicità dei batteri
Alcuni batteri sono parassiti di piante e animali, vale a dire vivono a loro spese, potendone causare o meno la morte.
Alcuni patogeni sono definiti opportunisti, in quanto sono generalmente innocui, ma possono arrecare danni considerevoli fino a provocare la morte nei soggetti con difese immunitarie deboli.
Molti patogeni opportunisti sono batteri normalmente presenti nell'ambiente o anche commensali dell'organismo e sono responsabili, ad esempio, della compromissione dello stato di salute dei soggetti ospedalizzati nei quali provocano infezioni spesso intrattabili con terapia antibiotica perché multiresistenti.
La patogenicità è definita come la capacità del batterio di causare la morte o anche lesioni progressive.
Il grado di patogenicità è detto virulenza.
Patogenicità e virulenza dipendono da proprietà strutturali e biochimiche riconducibili essenzialmente a tre meccanismi: invasione e colonizzazione dell'organismo ospite, capacità di resistere o addirittura contrastare le difese immunitarie, produzione di tossine.
I Gram positivi generalmente producono tossine di natura proteica liberate nell'ambiente esterno e dette esotossine.
Ne sono esempi la tossina botulinica e la tossina tetanica, prodotti rispettivamente da Clostridium botulinum e Clostridium tetani.
Le proprietà tossiche di alcuni Gram negativi dipendono da componenti dell'LPS.
Tra le malattie batteriche più comuni nell'uomo si ricordano: difterite, tubercolosi, polmonite, tetano, botulino, sifilide, gonorrea, gastroenteriti, meningite (causata da Neisseria meningitidis, ma anche da altri batteri), colera.
Link correlati:
Qual è la differenza tra virus e batteri?
Cosa sono i ferrobatteri?
Cosa sono le mutazioni geniche?
Che cosa sono le cellule natural killer?
Qual è la differenza tra epidemia e pandemia?
Studia con noi